肠道健康资讯
国际斑马鱼研究 | 益生元和益生菌对斑马鱼微生物群和肠道免疫的影响
2021-08-16 17:49 来源:新营养

文丨Adrià López Nadal ,Wakako Ikeda-Ohtsubo, Detmer Sipkema, David Peggs ,Charles McGurk, Maria Forlenza,Geert F. Wiegertjes and Sylvia Brugman 翻译丨周佳丽 DOI丨Frontiers in Immunology,February 2020 | Volume 11 | Article 114 www.frontiersin.org/10.3389/fifimmu.2020.00114
核心摘要 :使用斑马鱼作为实验模型引起了人们的兴趣,尤其是开发益生元和益生菌作为免疫和微生物组调节剂。尽管大多数益生元和益生菌都能确保对宿主有益,但对其影响的仔细评估仍然很重要,如人类益生菌的影响所显示的发现有问题的研究设计、报告不完整、缺乏透明度或安全性报告不足等。 在这篇综述中,我们首先描述了斑马鱼肠道的解剖结构和功能,然后,我们总结并批判性地回顾了最近关于益生元和益生菌与斑马鱼微生物群和免疫反应改变相关的大部分研究。 一、引言 1960 年代后期,匈牙利分子生物学家 George Streisinger 获得斑马鱼,以研究在脊椎动物模型中应用正向遗传学的分子机制。 最初,研究人员使用斑马鱼研究发育生物学,随后将斑马鱼应用于许多其他领域。其中,由于斑马鱼基因组中人类直系同源基因的高度存在(约87%)及其内在特征,斑马鱼作为研究免疫的模型脱颖而出。 与小鼠相比,斑马鱼体型小(<5 cm)、高产(每周200-300个新后代)且生长速度快。斑马鱼在子宫外发育,结合胚胎的透明度,可以研究个体发育体内从开发的早期时间点开始。 此外,使用转基因鱼有助于在体内显示特定的免疫细胞群,如中性粒细胞,这是基于中性粒细胞相关酶的荧光表达。它们注释良好的基因组简化了突变斑马鱼系的产生,其中一些有助于阐明免疫基因功能。 在过去十年中,基于Zinc finger nuclease、TALENs和非常成功的CRISPR-Cas的基因组编辑技术改变了在这种模式生物中可以解决单基因功能的速度。目前基因插入似乎仍然比基因敲除更具挑战性,这在不久的将来无疑会发生变化。斑马鱼的特征与这些独特的研究工具相结合,将这些斑马鱼确立为研究免疫过程和潜在分子机制的重要动物模型。 二、斑马鱼肠道:结构、功能和微生物群 斑马鱼没有胃,它们的消化道在解剖学上分为不同的部分:口腔、食道、三个肠段(前、中和后)和肛门。斑马鱼的食道与前肠段相连,其中,营养吸收主要是由于消化酶的含量高。营养吸收从前肠段到后肠段逐渐减少。离子转运、水重吸收、发酵过程以及某些免疫功能发生在肠中段和后段。 通过研究成年斑马鱼肠道的基因表达,并将其与小鼠的肠道进行比较发现,所有脊椎动物IEC中都存在的一组核心基因,以及转录起始位点和调控区域的保守性,与序列相似性无关。 除了上述所有相似之处,斑马鱼和小鼠消化道之间存在明显的解剖学差异。斑马鱼没有胃、肠隐窝、pp淋巴结和Paneth cells。此外,饮食习惯、环境条件、体型和或特定代谢要求也存在差异。 早期的定植研究表明,每个微生物的迁入和肠道适应都具有时间特异性:γ-变形菌在环境样本和斑马鱼幼鱼的肠道中含量很高,而β-变形菌在环境样品中含量最高。样本和幼年斑马鱼的肠道,表明在初次接触后某些种类的β-变形菌延迟定植。 斑马鱼幼体具有功能完善的器官,但其免疫系统尚未完全成熟。斑马鱼的适应性免疫成熟是科学领域内一个活跃的研究课题。在一项相对较小的研究中,我们发现T细胞控制斑马鱼肠道中变形菌的丰度,这提供了证据,证明与小鼠一样,适应性免疫系统在塑造微生物群组成方面发挥作用。如使用CD4-1:mCherry转基因斑马鱼和CD8a+抗体染色所示,T细胞以4 dpf存在于胸腺中。结果表明,T细胞早在10 dpf就从胸腺中排出。这表明从那个时间点开始,斑马鱼可能会产生系统性适应性反应。 在最初的定植期之后,对于宿主和微生物的发育都很重要,微生物群被认为进入稳定状态。野生捕获的斑马鱼和在两个独立实验室设施中饲养的斑马鱼的肠道微生物群的比较表明,存在一个共享的所谓核心肠道微生物群。高质量的16S rRNA基因分析显示常见和丰富的细菌群由21个操作分类单位(OTU)代表,主要是变形杆菌门(气单胞菌属和希瓦氏菌属),其次是梭杆菌或厚壁菌门(杆菌类)、放线菌门和拟杆菌门。 总之,地球上的所有生物都被其环境中的细菌物种定殖。宿主和定植微生物进行适应以确保宿主和微生物群的健康。斑马鱼可以饲养GF并且在10 dpf时仍然是光学透明的,加上免疫细胞群转基因的可能性,这一事实使斑马鱼成为研究微生物定植和免疫系统成熟时间的非常强大的生物。 三、益生元 益生元可定义为不可消化的成分,通过选择性刺激共生肠道细菌的生长或活性,从而改善宿主健康,对宿主产生有益影响。 益生元通常由小碳水化合物链组成,这些小碳水化合物链可作为葡萄糖(如β-葡聚糖)、半乳糖、果糖或甘露糖的低聚糖在市场上买到。我们总结了这些研究表格1,大多数研究是在胚胎斑马鱼中进行的,只有很少的研究是在成年斑马鱼中进行的。 斑马鱼研究中最常用的益生元是褐藻糖胶(主要存在于褐藻和褐海藻中的硫酸化多糖)、β-葡聚糖(从细菌和真菌的细胞壁中提取的β-D-葡萄糖多糖),有时还有其他的,例如低聚半乳糖。值得注意的是,关于益生元对微生物群的调节知之甚少,因为大多数审查的研究仅调查了它们的免疫刺激作用。
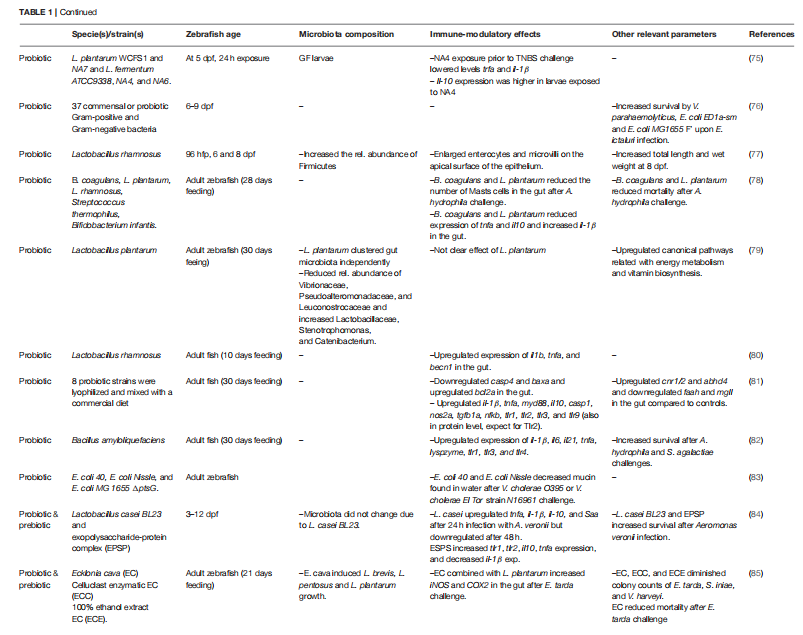
表格1 在斑马鱼中进行的关于免疫和微生物群的益生元、益生菌和合生元研究总结
从几种褐藻中提取的褐藻糖胶;将Eklonia cava ( 64 )、Chnoospora minima和Turbinaria ornata施用于水中的斑马鱼幼鱼。在所有三项研究中,暴露于岩藻多糖的幼鱼在LPS攻击后表现出活性氧(ROS)、诱导型一氧化氮合酶(iNOS)水平降低和整个幼鱼细胞活力提高。 然而,在这些研究中,当胚胎在受精后8 hpf时,候选益生元在水中被稀释。由于斑马鱼胚胎的在3 dpf才开口,完整的消化道要到6 dpf,此类研究并未证明益生元对肠道免疫有影响。优选地,具有完全发育的消化道(6 dpf或以上)的斑马鱼幼鱼被用于研究这种相互作用。 此外,在斑马鱼幼鱼中测试益生元可能会发现益生元功能,但总体目标通常是制定含有最佳益生元浓度的新型饮食。因此,幼鱼或成年斑马鱼更合适。我们研究了来自褐藻Cladosiphon okamuranus的岩藻依聚糖对整个斑马鱼幼鱼(水暴露)和成年斑马鱼肠道(用薄片喂养)中微生物群组成的影响。在成年斑马的肠道中,il-1β的基因表达减少,显性大肠杆菌(变形杆菌)减少,支持根瘤菌科和伯克霍德菌科,而在幼鱼il-1β、il-10、tnfb和mmp9增加,但没有观察到微生物变化。 与岩藻多糖不同,β-葡聚糖可以作为斑马鱼的免疫刺激剂。来自燕麦的β-葡聚糖,在5 dpf的全胚胎斑马鱼中以剂量依赖性方式上调了tnfa、il-1β、il-10、il-12、defb1、lyz和c-rel 的基因表达。在一项类似研究中,从4 hpf到6 dpf的β-葡聚糖暴露上调了tnfa、mpo、tlf和lyz基因表达。在这两项研究中,水中β-葡聚糖的给药阻碍了鱼对其摄取的量化,而且非常年轻的胚胎斑马鱼的暴露可能不会导致与肠道相关的影响。低聚糖如低聚半乳糖(GOS)和低聚果糖(FOS)经常被用作农业和人类婴儿营养中的益生元,通过增加建议的有益细菌发酵产物的产量来促进健康。以0.5%、1%和2%的包含水平喂食GOS 8周的成年斑马鱼显示出tnfa和lyz表达的上调以及整个斑马鱼中总免疫球蛋白的增加。然而,没有评估肠道特异性读数。 很明显,益生元可以根据其来源以特定方式作用于免疫系统。岩藻多糖可以减少炎症标志物,而β-葡聚糖和GOS会增加促炎细胞因子的基因表达。尽管取得了有希望的结果,但绝大多数研究将未发育的胚胎斑马鱼暴露于无法通过自由喂养摄取添加剂的益生元。如果将此类候选益生元包含在干颗粒中,并在饱食前稍稍给鱼服用(确保鱼吃掉所有颗粒),则可以更清晰地估计益生元肠道水平并评估对肠道微生物群和免疫的影响。 前面提到的益生元研究中尚未广泛采用的几种方法也可能适用于斑马鱼的益生元肠道健康研究。首先,需要组织学和免疫组织化学染色来了解肠道组织中的免疫调节作用(即正常肠道结构的破坏)。转基因斑马鱼可能有助于使用荧光激活细胞分选和成像来阐明哪些免疫细胞亚群渗入肠道。此外,这些亚群的细胞分选与转录组学将描述益生元的真正作用。 四、益生菌 早在1907年,Elie Metchnikoff就将益生菌的使用与预期寿命的延长联系起来。出于本综述的目的,我们将益生菌定义为活的或灭活的微生物,例如细菌或酵母菌,当通过水给予它们时,可以为宿主带来益处,例如提高抗病性或增强免疫反应。 益生菌可以通过多种方式影响宿主的健康:分泌抑制微生物病原体生长的次级代谢物或直接刺激免疫反应以下调肠道炎症。在这里,我们重点研究了斑马鱼中有关(肠道)免疫和微生物群调节的益生菌研究(总结于表格1)。 为了评估活益生菌的潜在健康益处,了解它们在宿主体内的最佳环境(氧气水平、pH值等)及其定植途径非常重要。益生菌-宿主相互作用通过GF斑马鱼口腔-肠道病原体定植模型解决。 首先,将6 dpf斑马鱼浸泡在25种潜在的肠道鱼类病原体中,然后在3天内记录死亡率。Edwardsiella ictaluri引起的幼鱼死亡率最高,并被进一步选择来挑战这种鱼。 然后,幼鱼在E. ictaluri攻击之前用37种可能的益生菌的单一菌株进行预定殖。通过这种广泛的筛选,副溶血性弧菌、大肠杆菌ED1a-sm和E.coli MG1655 F'显着提高了E. ictaluri感染后的存活率。V. parahaemolyticus通过抑制E. ictaluri的生长来保护宿主,而大肠杆菌则通过特定的粘附因子来保护宿主,例如F pili参与生物膜和结合形成,为宿主中的其他益生菌提供生态位。 值得注意的是,斑马鱼的鳃虽然在受精后2周内活跃于气体交换中,但却为病原体提供了一个潜在的入口。有趣的是,在同一项研究中,副溶血性弧菌被评估为可能的益生菌,而鱼肠弧菌被认为是可能的病原体。大多数微生物群研究将免疫反应与分类学水平相关联,例如属或科(即弧菌属),而不是物种或菌株。因此,将整个属推广到可能是物种甚至菌株特异性的功能。这种广泛使用的概括可能来自难以生成足够长的扩增子以区分密切相关的生物体。 此外,转录组学和散弹枪方法优于16S rRNA基因分析来描述活跃的微生物群,因为它们提供更多关于鱼类健康状况的信息。成年斑马鱼也被用来测试益生菌作为人类益生菌消耗的模型。 将成年斑马鱼暴露于两种大肠杆菌菌株(Nissle和MG 1655 ΔptsG),并用霍乱弧菌(El Tor 菌株)进行攻击。大肠杆菌属减少水箱水中的粘蛋白含量,这是腹泻的指标,尽管这些粘蛋白也可能是由于皮肤脱落造成的。评估这些大肠杆菌是否存在可能会很有趣。如前所述,通过减少Myd88-Notch信号来增加分泌细胞的发育,从而增加粘液分泌。 此外,虽然在人类中通过口服溶液给予细菌是确保摄入的有效方法,但向水中添加益生菌可能无法保证被鱼吸收,并可能影响整个鱼的粘膜(皮肤、鳃、肠道),不仅被肠道吸收。鱼肠道的环境比人类肠道环境更需氧,在这些需氧条件下,乳酸菌可能会被其他细菌击败。这个基本原理可以解释为什么通过浸泡在斑马鱼中测试的人类益生菌(乳酸杆菌属)不能提供对E. ictaluri感染的保护。 几项研究报告称,乳酸菌(LAB)是良好的益生菌候选物,因为它们能够耐受和粘附在肠道上,产生的乳酸可抑制病原菌的生长,并加强粘膜屏障。在用嗜水气单胞菌进行浸泡挑战后,与对照相比,从3-25 dpf浸入干酪乳杆菌BL23的斑马鱼显示出更高的存活率。与对照相比,在嗜水空气单胞菌攻击后8和24小时后,il-1β、C3a和il-10的肠道基因表达上调。 有趣的是,来自该属的潜在益生菌乳酸杆菌以菌株特异性方式调节基因调控。事实上,与对照相比,浸入发酵乳杆菌NA4的GF幼鱼在化学诱导的炎症后显示出增加的il-10表达和减少的il-1β和tnfa表达。 然而,在同一项研究中,斑马鱼幼鱼浸泡在几种植物乳杆菌菌株(WCFS1和NA7)或其他发酵乳杆菌菌株(ATCC9338和NA6)中并没有表现出这些基因表达差异。上述研究中基因表达的差可能是由于鱼龄、组织分析(肠道与整个幼鱼)所应用的挑战(活病原体与化学物质)以及用作益生菌候选物的特定乳杆菌菌株。 淀粉芽孢杆菌在商业饮食中每天补充两次,持续30天,上调il-1β、il-6、il-21tnfa、溶菌酶、tlr1、tlr3。尽管没有考虑其他免疫参数,但基因表达的上调似乎与增强的先天免疫有关。 在另一项针对成年斑马鱼的研究中,商业饮食补充了多种冻干益生菌菌株30天。益生菌混合物上调了肠道中il-1β、tnfa、myd88、il-10、casp1、nos2a、tgfb1a、nfkb、tlr1、tlr2、tlr3和tlr9的表达。 此外,益生菌混合物增加了所有上调基因(Tlr2蛋白除外)编码的蛋白质水平。一方面,益生菌混合物中的某些细菌可能抑制了Tlr2,这反过来又可能部分抑制了myd88。另一方面,益生菌混合物中的其他细菌可能增强了其他TLR的表达,这些TLR上调了myd88和整体Myd88平衡协调的先天免疫反应。 如前所述,微生物物种可以影响宿主免疫力,而不管它们的丰度如何,并且当混合使用益生菌时,每个物种的影响更难解开。其他使用LAB作为益生菌的研究不仅检测了基因表达,还检测了斑马鱼肠道中的微生物群和组织学变化。 一些研究调查了酵母作为斑马鱼益生菌的潜力。GF和CONV斑马鱼幼鱼从2-3 dpf浸入两种酵母的溶液中,然后在14 dpf时对肠道微生物群进行采样。尽管观察到微生物变化,但未测量免疫相关结果,因此本研究中酵母菌的益生菌作用仍未确定。 在另一项研究中,4 dpf斑马鱼在受到攻击前暴露于15种荧光标记的酵母菌株2小时。大多数酵母菌株在攻击后具有增加的存活率。在后来的实验中,同一组使用类似的设置进一步研究了GF和CONV幼鱼中的两种酵母菌株。在攻击后,暴露于任一酵母菌株显着增加了GF和CONV幼鱼的存活率。被攻击的CONV斑马鱼表现出il-1β、c3、tnfa、mpx和il-10表达的上调。用任一酵母菌株进行预处理可阻止CONV和GF幼鱼中的此类基因上调,表明这些酵母菌株可能会阻止或降低攻击的影响。 斑马鱼也被用于合生元研究,这些研究通常结合使用益生元和益生菌。干酪乳杆菌BL23和胞外多糖复合物(ESPS)在3至12 dpf的GF和CONV幼鱼中进行了组合研究。干酪乳杆菌暴露于维罗单胞菌攻击24小时后上调tnfa、il-1β、il-10和saa表达,并在攻击48小时后下调这些基因的表达。 值得注意的是,在24小时攻击后,单独的ESPS上调tlr1、tlr2、il-10和tnfa并下调il-1β。合生元,干酪乳杆菌BL23和EPSP在A. veronii攻击后剂量依赖性地提高存活率。将具有增强生物活性的E. cava酶消化物作为益生元与作为益生菌的植物乳杆菌在成年斑马鱼中联合补充21天,可降低肠道中iNOS和环氧合酶2 ( cox2 )的水平。此外,当益生元和益生菌一起给药时,与单独用植物乳杆菌处理的鱼相比,它们在用迟发性大肠杆菌攻击后增加了存活率。有趣的是,这些研究表明某些提取物或生物活性化合物而不是整个益生元可能会引起免疫调节。 大量研究将潜在的益生元和益生菌暴露于斑马鱼,以通过微生物调节来改善它们的免疫状况(图1)。
图1 斑马鱼肠道中益生元和益生菌、免疫系统和微生物群相互作用的概述
值得注意的是,在大多数这些研究中,基因表达被认为是决定性的免疫读数。除了基因表达并不总是转化为蛋白质功能这一事实外,促炎和抗炎细胞因子通常根据反应的动力学和时间而上调或下调。基因表达可能反映免疫反应期间宿主的平衡:特异性和强度足以对抗潜在的致病细菌,但同时能够耐受共生宿主微生物群。这种平衡还取决于不同的细胞类型,它们协同工作以防止在对抗入侵病原体或持续炎症时对宿主造成过度损害。 我们需要更详细地了解参与不同反应的不同免疫细胞类型的作用和存在,然后才能尝试调节对宿主有益的反应。为此,斑马鱼仍然是理想的候选模式生物。迄今为止,更多的研究可以利用斑马鱼的独特工具,例如不同转基因报告斑马鱼(细胞因子以及免疫细胞群)的实时成像,以更广泛地了解宿主-微生物的复杂动态相互作用相互作用。 五、结论 最近,使用小鼠模型进行的新研究表明,微生物群落可以影响病毒感染的严重程度。此外,使用RAW264.7细胞的体外数据显示,几种乳杆菌菌株通过IFN-β上调对鼠诺如病毒(MNV)感染具有抗病毒活性。目前,尚不清楚微生物是否也能改变鱼类特有的病毒感染性。这是一个令人兴奋的新研究途径,可能会导致新的疫苗接种策略,将靶向病毒的疫苗与益生元或益生菌治疗相结合,以改变微生物群并靶向病毒本身。由于其独特的优势,斑马鱼最有可能成为其中的一个基本领域。 使用斑马鱼在该领域发表的研究将继续增加,并结合现有技术(组学、免疫组织化学、FACS、体内成像)或通过新兴的新技术知识空白肯定会被填补。对于未来的实验,如果采用更全面的方法,将大大有助于我们的理解。我们需要在每个实验中结合读取参数,例如基因表达、攻击后存活率、肠道结构、免疫细胞募集、微生物群组成、代谢物产生和行为数据,以提供某些治疗对健康的后果的更广泛的了解。只有通过基因编辑询问可能的分子途径来仔细确定因果关系,我们才能为新型免疫调节策略的设计提供坚实的理论基础。新营养留言互动
您的电话不会被公开。 必填项已用 * 标注
相关热词搜索:
上一篇:打造朋克养生爆品靠类生元®(吉品后生元/灭活菌): 「L-137热封型乳酸菌」&「LM-1004享受乳酸菌」
下一篇:从新研究、新功能、新概念看牛初乳华丽变身 | 营养盒子直播
新营养峰会/活动


新营养热门资讯

共晶技术:让营养素步入埃米时代
XINGRAPHIC · 09-11

11月6-7日广州新营养趋势大会!30+演讲、2
风信社 · 09-08

当“造物”成为可能,人类营养健康产业将迎
XINGRAPHIC · 07-07

新营养周报 | 农夫山泉上新蓝靛果混合汁
数据 / 观察 · 01-06
新营养合作伙伴